Le premier chercheur ayant utilisé de véritables techniques
génétiques fut Gregor Mendel vers1856 et le résultat
est remarquable (sinon remarqué àl'époque) puisqu'il proposa une "théorie
particulaire del'hérédité" pleinement confirmée par ladémonstration récente
selon laquelle l'informationgénétique est codée d'une façon discontinue,en
gènes, dans la molécule d'ADN.
Il n'est pas question ici de retracer toute l'oeuvre de Mendel ni de consacrer
une part importante de ce cours à la génétique formellemais la méthode d'analyse
mendélienne est d'autant plusd'actualité que l'étude du polymorphisme de l'ADN
telle qu'onla pratique maintenant fait appel aux mêmes principes.
De très nombreux biologistes avant Mendel avaient observé lerésultat de croisements
de plantes ou d'animaux sans en tirer le moindreenseignement. Pourquoi Mendel
a-t-il compris l'essentiel du mécanismede l'hérédité en réalisant simplement
des croisementsde pois et en n'observant que quelques caractères
phénotypiques ?
La démarche sert de modèle de nos jours.
- il a choisi, comme matériel d'étude, une espèce à cycles relativement courts, autogame (dont les fleurs s'autofécondent) mais malgré tout à fleur accessible et pollinisable manuellement.
- il existait de nombreuses variétés de pois aisément discernables les unes des autres.
- il s'est intéressé à des caractères qualitatifs bien tranchés : aspect lisse ou ridé du grain, couleur de la fleur, des cotylédons ... dont l'expression n'est pas soumise à l'environnement. Les caractères quantitatifs, qui peuvent varier d'une façon continue (taille de la plante, poids des grains ...) sont beaucoup plus difficiles à appréhender bien que se transmettant selon les mêmes règles que les autres.
- il n'a étudié que peu de caractères à la fois (un seul au début).
- il a été le premier à utiliser l'outil mathématique en biologie.
- il avait un grand sens de l'analyse et de la synthèse et ne s'enlisait pas dans les détails.
1 ANALYSE MENDELIENNE DE LA TRANSMISSION D'UN CARACTERE
(MONOHYBRIDISME)
Elle va se faire par croisement de variétés ne différant que
par un caractère d'où le terme de monohybridisme souvent utilisé.
L'exemple le plus célèbre est celui du caractère deforme de la graine de pois,
ronde chez la plupart des pois mais ridéepour plusieurs variétés établies
à l'époquede Mendel. "Variété établie" veut dire ici (et ce futl'une des expériences
de Mendel) que les graines produites par desplantes issues de graines rondes,
par autofécondation, présentent la même caractéristique de productionde graines
rondes. Il en est exactement de même pour des "lignées"à grains ridés.
C'est cette notion de continuité génétique qui permet l'obtention de ce que nous allons appeler des lignées pures.
Ayant observé et sélectionné ce caractère nettementdiscriminant,
il va croiser des plantes de ces deux lignées, plantesdéfinies comme parents
(P). Le résultat est l'obtention deplantes (de première génération, appelée
F1)qui toutes produisent des grains ronds. L'homogénéitédes produits de première
génération est remarquableet va constituer un élément important de l'analyse.
Unegénération F2 obtenue par autofécondation d'individusF1 présente à nouveau
les deux phénotypes : grains rondset ridés, le dénombrement fait ressortir
dans cette populationde seconde génération trois quarts de grains ronds pour
unquart de grains ridés : les deux caractères parentaux serépartissent (ségrégent)dans
des proportions 3/4, 1/4.
Mendel a pu généraliser cette observation en croisant d'autreslignées ne différant
que par un caractère, 7 "traits"qualitatifs ont été suivis avec les résultats
suivants:
| Lignées parentales | Phénotype
F1 |
Rapport phénotypique
des plantes F2 |
| grains ronds x ridés grains jaunes x verts pétales pourpres x blancs gousses pleines x plissées fleurs axiales x terminales tiges longues x naines |
ronds jaunes pourpres pleines axiales longues |
2,96 ronds pour 1 ridé 3,01 jaunes/1 vert 3,15 pourpres/1 blanc 2,95 pleines/1 plissée 3,14 axiales/1 terminale 2,84 longues/1naine |
L'exemple de la couleur de la fleur a étéchoisi dans le cours illustré.
L'observation des produits de l'autofécondation de lagénération
F2 était la suite logique du travail. Lerésultat pour le caractère "aspect
du grain", a étéle suivant : les plantes issues des pois à phénotype grainridé
reproduisent invariablement le même phénotype. Ilen va autrement des pois
de phénotype grain rond, bien que certainsreproduisent invariablement le caractère
grain rond, d'autres produisentun mélange des deux phénotypes dans des proportions
de 3/4de ronds pour 1/4 de ridés.
Le côté pratique des expériences, regroupées dansla figure 1.1 sont à la portée
de tout le monde, encore fallait-illes interpréter.
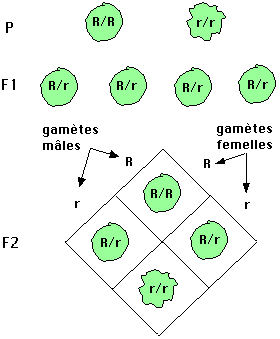
Il faut procéder méthodiquement en tirant parti de l'aspect qualitatif puis quantitatif des résultats.
Sur le plan qualitatif, il est clair qu'uneinformation est transmise et non un caractère: aucune plante ne présente le caractère ridé en F1cependant il réapparait en F2. Il faut bien admettre que ces informationssont conservées intactes (pures) à travers lesgénérations. Toute plante possède en fait deux informationspour un même caractère (deux formes alléliques), l'uned'entre elles (ridé pour la forme du grain ou blanc pour la couleurde la fleur...) n'étant pas exprimée en F1, on dira quel'allèle (rond ou pourpre) est dominantsur l'autre. L'allèle masqué en F1 est appelérécessif. Mendel a perçu ladiploïdie de la cellule somatique et l'asymbolisé par des lettres :
R (majuscule) va représenter l'allèle rond dominant, r (minuscule),l'allèle ridé, récessif.
L'information (double) pour une plante de lignée pure de phénotypegrain
rond sera écrite R/R c'est ce qui représente le"génotype"
de la plante pour lecaractère forme du grain. De la même façon, le génotypede
la lignée récessive pour ce même caractèresera écrit r/r .
Mendel a également compris qu'au cours de la formation des gamètesil y a disjonction
de ces deux informations,ce que l'on connaît actuellement de la méiose
et de laséparation des chromosomes homologues (porteurs d'élémentsd'information
homologues), apporte un support à l'hypothèsede Mendel.
Il est clair qu'une recombinaison va avoir lieu au moment de lafécondation,
les combinaisons parentales d'information sont :
R avec R pour l'un avec production de gamètes R et r avec r pour l'autreet
production de gamètes r, l'information d'un individu issu de cecroisement
regroupera R et r et le génotype s'écrira R/r. Cecireprésente une combinaison
nouvelle par rapport à R/R ou r/ret constitue une première prise de contact
avec la notion fondamentaleen génétique de recombinaison("Re
- combinaison").
Un pas considérable sera franchi (bien après Mendel) lorsquel'on
découvrira que derrière ces majuscules et minuscules (Ret r par exemple),
qui symbolisent des formes alléliques d'un mêmegène, se cachent des séquences
d'ADN, presque semblables, mais pas tout à fait, situées au même endroit (au
même locus), dans deux chromosomeshomologuesDe
la même façon, les réponses à la question de l'expression du matériel génétique
vont éclairer des notions restées longtemps abstraites telle que la dominance
parexemple.
Sans attendre le détail des chapitres relatifs à l'expression des gènes, il
est bon d'avoir présent à l'esprit, d'une façon réflexe, quel'expression d'un
gène passe par la réalisation d'une protéine spécifique, le
gène,en tant qu'unité d'information "code" une protéine.On peut en
rester là : un caractère phénotypique peuttrès bien être une protéine de structure.
Il peutégalement s'agir d'une protéine enzymatique et un exemple très simple
de la réalisation du caractèrephénotypique "fleur pourpre" étudié par
Mendel, pourraitêtre le suivant :
![]()
L'enzyme Enz représente le produit direct d'un gène.
En reprenant la symbolique déjà utilisée, si E représente l'information pour Enz :
E/E représente un génotype homozygote avec, sur chaque chromosome homologue une information permettant la synthèse de l'enzyme fonctionnelle et donc la possibilité de transformer le substrat incolore en pigment.e/e représente un génotype, homozygote également mais,pour chaque chromosome homologue, le locus est occupé par une information erronée ne permettant pas la synthèse d'une enzyme fonctionnelle, en conséquence, le substrat ne sera pas métabolisé et la fleur apparaître blanche (absence de pigment).L'hétérozygote E/e possèdeà la fois une information normale et une information erronée,l'information correcte permet la réalisation d'une enzyme fonctionnelleet la synthèse du pigment car elle"complémente" l'information incorrectece qui se traduit par : l'allèle E est dominant sur l'allèlee (récessif) car chez un hétérozygote E/e c'est lephénotype pigmenté qui se manifeste.
Cette incursion dans le domaine de l'expression de l'informationgénétique va nous permettre d'aborder, à l'aide d'unseul exemple, deux notions importantes (par leurs conséquences), lacodominance et le pluriallélisme.
2 CODOMINANCE ET PLURIALLELISME
Prenons un exemple en génétique humaine. Les groupes sanguins
sont définis par la présence d'antigènes spécifiques à la surface des hématies.
Dans le système ABO, nous allons nous intéresser aux antigènes "A" et "B".
La production de ces molécules dépend d'informations génétiques situées à
un locus précis d'un chromosome (I). Ce locus peut porter le code de l'enzyme
responsable de la synthèse de l'antigène A et serasymbolisé par IA,
l'individu de génotype IA/IA
(homozygote) va donc synthétiser des antigènes de type A, l'individu IB/IB
portel'information pour des antigènes de type B et va les produire. Un hétérozygote
IA /IB possède deux
informations différentes correspondant à deux allèles différents pour un
même locus, le
phénotype de cet individu est AB, les deux informations sont exprimées (les
deux antigènesA et B sont présents), on dit qu'il y a
codominance.
Dans une cellule diploïde, il est évident que deux allèles seulement peuvent
coexister car il n'existe que deux chromosomes homologues,ceci ne veut pas
dire qu'il n'existe que deux allèles possibles capablesd'occuper ce locus,
on verra que théoriquement il peut en apparaître une infinité. Dans le cas
des groupes sanguins, il existe un allèle qui ne spécifie aucune enzyme capable
de produire un antigène (la fonction est perdue, allèle nul), on symbolise
cet allèle par i .
En conséquence, un individu IA /i portera une information
"antigène A" sur un chromosome et une information "aucun antigène" sur le
chromosomehomologue (le fait qu'une mutation conduise à un allèle "nul"sera
expliqué au chapitre V) , pour l'instant, ceci nous permet decomprendre que
l'individu sera du groupe A (on dira que IA est
dominant) de la même façon que l'individu de génotype IB/i
serade groupe B, seul un homozygote récessif i/i sera
de groupe 0 (avant l'informatique on ne distinguait
pas la lettre O du zéro).
La suite du travail de Mendel, l'analyse d'autres variants, a permis d'aboutirà une notion de gènes en tant qu'unités d'informationsdiscontinues et indépendantes. Il s'agit essentiellement de l'étude de variants se distinguant par deux, trois ou plusieurs caractère(cas de di, tri, et polyhybridisme).
3 DIHIBRYDISME
Comme on l'a vu, Mendel a montré expérimentalement que lesmêmes règles régissent la transmission de 7 caractères phénotypiques chez le pois. Or, s'il envisage des variétés différant par deux de ces caractères, par exemple une àgrains ronds de couleur jaune et l'autre avec des grains ridés etverts, le croisement de ces lignées parentales donne une F1 homogèneavec des grains ronds et jaunes (ceci confirme la dominance de ces caractères), et une F2 dont les phénotypes sont les suivants:
rond - jaune : 9/16ème de la population
ridé - vert : 1/16ème
rond - vert : 3/16ème
ridé - jaune : 3/16ème
Pour les deux caractères pris séparément, les proportionsde phénotypes sont 12/16ème et 4/16ème soit les classiques3/4 et 1/4, les gènes se comportent donc d'une façonindépendante, les unités d'information pour différentscaractères sont séparées les unes des autres : on diraqu'il y a ségrégation indépendante de ces unités à la méioseet recombinaison à la fécondation comme le fait apparaîtrel'analyse des génotypes :
| rond jaune ridé vert | |
| types parentaux : F1 : autofécondation ségrégation indépendante |
R/R J/J
x
r/r j/j R/r , J/j (rond, jaune) R/r J/j x R/r J/j R,J et r,j de types parentaux |
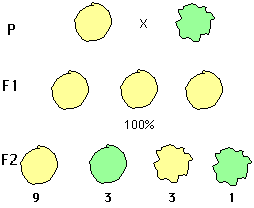
Figure 1.2
La rencontre au hasard de ces gamètes et le phénomènede dominance produira une population F2 telle que l'a observée Mendel.
4 GENES ET CHROMOSOMES
Il faudra attendre la reprise des travaux de Mendel par le groupe de Morgan, au début du siècle, travaillant sur un autre modèle,la drosophile, pour :
- confirmer les déductions de Mendel quant à la notion de gène et au mode général de transmission de ces éléments d'information
- interpréter (sinon découvrir) une exception de taille : les gènes ne se transmettent pas toujours indépendamment les uns des autres.
La prédiction des génotypes dansla figure 1.2 est de 50% de types parentaux et 50% de types recombinés,en étudiant le di ou trihybridisme chez la drosophile (il existe un nombre impressionnant de variants) on peut se rendre compte que certainsde ces caractères ne répondent pas à cette prédictionet semblent se transmettre "en bloc", c'est à dire que la F2 fera apparaître beaucoup plus d'associations parentales que recombinées.
La "théorie chromosomique de l'hérédité" a pris jour en remarquant l'analogie entre le comportement des facteurs mendéliens et celui des chromosomes au moment de la méiose puis en constatant une liaison entre certains gènes et des chromosomes spécifiques, les chromosomes sexuels.
Sans le savoir, Mendel avait étudié, chez le pois, 7 caractères portés par 7 chromosomes différents !
En même temps que l'on découvrait la liaison de sériesde gènes sur les chromosomes, on constatait que cette liaison n'était pas absolue et qu'il existait une possibilité de recombinaison entre des gènes situés dans les deux chromosomes homologues (d'une même paire).
Malgré le parallèle cytologique avec les chiasmas, il faudra attendre de connaître la composition chimique des chromosomes et lastructure de l'ADN pour bien comprendre les mécanismes du "crossingover" soit de la recombinaison à l'échelon moléculaire.Quoi qu'il en soit, dès Morgan, l'analyse des recombinants chez drosophile a permis l'apparition des premières bases de la cartographie génétique.
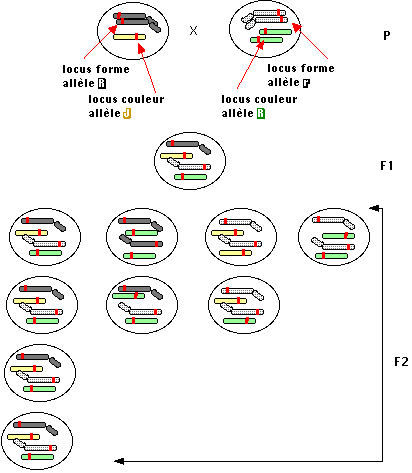
Figure 1.3 : Interprétation chromosomique de la figure 1.2
| Le génotype est un ensemble
d'informations discontinues : les gènes. Ceux-ci sont organisés de façon
linéaire en groupes de liaisons supportés physiquement par le chromosome.
Le degré variable de liaison entre deux gènes détecté par la fréquence
de recombinaison reflète la distance qui les sépare et permet de construire
des cartes génétiques. Les gènes ne sont détectables que parce qu'ils peuvent varier et exister sous plusieurs formes alléliques. |
Dans ce premier chapitre, le gène représente non seulementune unité de fonction mais également une unité derecombinaison et une unité de mutation, ceci ne va pas s'avérerexact.
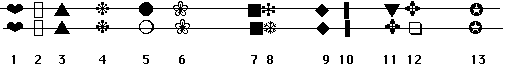
Représentation "en collier de perles" d'un génome diploide.
Les loci 5, 8 et 12 sont héterozygotes, les autres sont homozygotes.
| relire ? |
