Malgré cela, l'analyse, pourtant relativement simple, du dihybridisme (avec dominance) qui se traduit, en F2, par quatre phénotypes dans des proportions 9-1-3-3 alors qu'il existe neuf génotypes possibles, laisse présager que le petit pois et la drosophile sont des organismes (diploïdes) bien trop compliqués pour saisir les mécanismes fondamentaux de la recombinaison et de l'expression des gènes.
| Les génomes haploïdes tels que ceux des bactéries et des virus vont révolutionner la notion de gène. |
Des séries de travaux s'étalant de 1928 à 1952 vont permettre d'associer définitivement l'ADN à la notion d'information génétique. Ces travaux sont exposés ci dessous car ils présentent, comme ceux de Mendel, beaucoup plus qu'un intérêt historique : ils sont des modèles d'analyse faisant appel, pour la première fois, à des méthodes de la biologie moléculaire.
1.1 TRAVAUX DE GRIFFITH :
Le point de départ est une bactérie pathogène, un pneumocoque, agent de la pneumonie chez l'homme (et, au laboratoire, létal pour la souris). Cet exemple rappelle que ce sont les pathologistes qui ont amené les généticiens à étudier les procaryotes et les virus.
La virulence de bactéries représente déjà un caractère phénotypique, un autre est apporté par l'aspect des colonies qui se développent à la surface d'un milieu nutritif gélosé lorsque l'on a ensemencé une boite de Pétri : un mucopolysaccharide secrété par les bactéries donne un aspect huileux aux colonies, ce phénotype est baptisé "S" (comme smouth = lisse).
Lors de repiquages et d'étalement successifs, on voit parfois apparaître des colonies présentant un phénotype différent : "rugeux" par opposition à lisse désigné par "R" (comme rough).
Inoculées à une souris, les bactéries de type R s'avèrent non virulentes. Les deux phénotypes sont liés.
| * Remarque : L'apparition, rare et spontanée, de variants dans une population bactérienne est le résultat d'une mutation. A l'origine, une seule bactérie est modifiée mais cette modification étant héréditaire, la colonie que l'on observe représente en fait la descendance de la bactérie mutée. |
| * Remarque : Selon
des mécanismes qui seront détaillés plus tard, l'accident mutationnel
peut, dans certains cas, se produire "dans l'autre sens" : dans une
population R peut spontanément réapparaître un individu S, cette modification
est, elle aussi héréditaire puisque ce que l'on observe c'est une colonie
S c'est à dire une descendance. . Une des caractéristiques de ce phénomène de réversion est qu'il se produit avec la même fréquence que la mutation : c'est un événement très rare. Cette notion de réversion, simplement effleurée ici, s'avérera très importante. |
Vers 1928, Griffith réalise une expérience fondamentale :
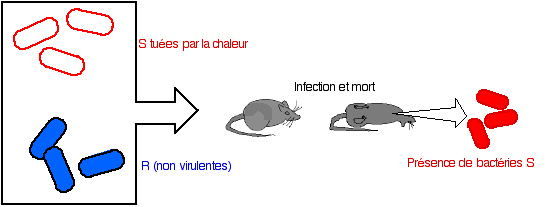
- première étape : il inocule à une souris, des bactéries S (de phénotype virulent) tuées par la chaleur, les souris ne présentent aucun trouble.
- deuxième étape : il inocule des bactéries S tuées par la chaleur après les avoir mélangées à des bactéries R (phénotype non virulent), des souris meurent de pneumonie. Le prélèvement de bactéries à partir de souris mortes et leur mise en culture révèle un phénotype S (et virulent) pour toute la descendance.
Interprétation : les bactéries vivantes au départ sont de type R, non virulent. Griffith, suppose qu'elles ont été "transformées" par un élément provenant des bactéries tuées par la chaleur. La fréquence de la transformation et des arguments d'ordre immunologique excluent un phénomène de réversion.
A l'époque, Griffith ne peut que conclure à l'existence d'un facteur transformant or il s'agit bien d'une transformation génétique au sens actuel du terme.
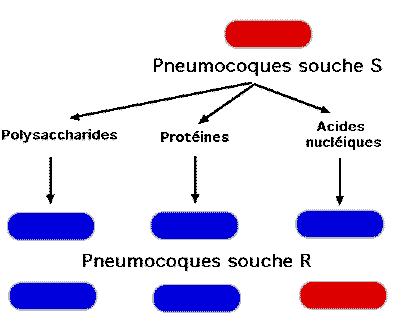 En
1944, Avery, Mc Leod, Mc Carthy reprennent ces travaux avec des tests un peu
plus sophistiqués qui préfigurent la génétique microbienne moderne : ils mélangent,
dans un tube, des bactéries S tuées par la chaleur et des bactéries R. Après
un temps de culture suffisamment long, un anticorps anti R est ajouté, (plusieurs
souches de ces bactéries ont été répertoriées ,qui diffèrent par leurs propriétés
antigéniques), les bactéries R sont agglutinées et sédimentent au fond du
tube, le surnageant est ensuite étalé sur un milieu nutritif et l'on s'aperçoit
que des colonies de phénotype S se développent. Cette manipulation in vitro
va servir à l'identification du principe transformant en ajoutant à des bactéries
R, non plus des S tuées par la chaleur mais des extraits relativement purifiés
de celles-ci. Les auteurs de ce travail se tournent d'abord vers les polysaccharides
de la paroi, sans aucun résultat puis vers les protéines sans plus de succès,
ils n'obtiennent quelques cas de transformation qu'avec des préparations d'acides
nucléiques et particulièrement d'ADN.
En
1944, Avery, Mc Leod, Mc Carthy reprennent ces travaux avec des tests un peu
plus sophistiqués qui préfigurent la génétique microbienne moderne : ils mélangent,
dans un tube, des bactéries S tuées par la chaleur et des bactéries R. Après
un temps de culture suffisamment long, un anticorps anti R est ajouté, (plusieurs
souches de ces bactéries ont été répertoriées ,qui diffèrent par leurs propriétés
antigéniques), les bactéries R sont agglutinées et sédimentent au fond du
tube, le surnageant est ensuite étalé sur un milieu nutritif et l'on s'aperçoit
que des colonies de phénotype S se développent. Cette manipulation in vitro
va servir à l'identification du principe transformant en ajoutant à des bactéries
R, non plus des S tuées par la chaleur mais des extraits relativement purifiés
de celles-ci. Les auteurs de ce travail se tournent d'abord vers les polysaccharides
de la paroi, sans aucun résultat puis vers les protéines sans plus de succès,
ils n'obtiennent quelques cas de transformation qu'avec des préparations d'acides
nucléiques et particulièrement d'ADN.
| Autrement dit, une bactérie peut acquérir de nouveaux caractères phénotypiques, de nouvelles fonctions métaboliques (sécrétion de polysaccharides, virulence) par l'intermédiaire d'ADN provenant d'une autre. |
L'importance extraordinaire de ces travaux n'a pas été reconnue pendant longtemps pour plusieurs raisons :
- la structure chimique de l'ADN bien que déterminée d'une façon très incomplète, semblait trop simple pour pouvoir contenir une information aussi complexe que l'information génétique. Les des protéines en faisait de bien meilleures candidates comme support de cette information.
- la génétique microbienne était à ses début et l'on n'était
pas certain que le passage d'un type S à un type R soit le fait de 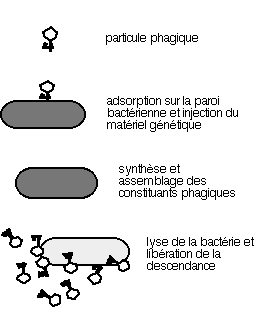 mutations,
ni que l'hérédité des organismes supérieurs soit comparable à celle des bactéries.
mutations,
ni que l'hérédité des organismes supérieurs soit comparable à celle des bactéries.
1.2 TRAVAUX DE HERSHEY ET CHASE :
En 1952, Hershey et Chase étudient la reproduction du bactériophage
T2 dans la bactérie Escherichia coli (qui deviendra le monstre sacré de la
génétique moderne) et emploient une technique qui va se développer rapidement
: l'utilisation d'isotopes radioactifs comme traceurs, comme marqueurs permettant
de suivre la destinée de macromolécules. Dans un premier temps, ils cultivent
les bactéries (E. coli) sur un milieu contenant du phosphore 32 (32P) et du
soufre 35 (35S), après un certain temps de culture, les éléments constitutifs
des bactéries contiennent ces marqueurs. On infecte alors la culture avec
une suspension de phages T2, ceux ci vont réaliser un cycle lytique en utilisant
les molécules radioactives de leurs cellules hôtes. La descendance phagique
est recueillie et sert à infecter des bactéries normales.
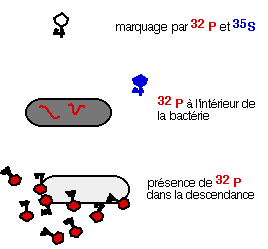 On sait que,
dans la première étape de l'infection, les phages s'adsorbent sur la bactérie
et injectent à l'intérieur une molécule informative. Après cette étape d'adsorption,
les auteurs agitent violemment la suspension pour décrocher ce qui reste à
l'extérieur des bactéries, après centrifugation, on obtient un culot bactérien
contenant l'information phagique et un surnageant contenant la capside (Figure).
Or, le culot contient le 32P et le surnageant le 35S, c'est la démonstration
éclatante que l'information génétique du bactériophage, qui pénètre à l'intérieur
de la bactérie est de l'ADN et que la capside protéique ne sert que d'emballage.
On sait que,
dans la première étape de l'infection, les phages s'adsorbent sur la bactérie
et injectent à l'intérieur une molécule informative. Après cette étape d'adsorption,
les auteurs agitent violemment la suspension pour décrocher ce qui reste à
l'extérieur des bactéries, après centrifugation, on obtient un culot bactérien
contenant l'information phagique et un surnageant contenant la capside (Figure).
Or, le culot contient le 32P et le surnageant le 35S, c'est la démonstration
éclatante que l'information génétique du bactériophage, qui pénètre à l'intérieur
de la bactérie est de l'ADN et que la capside protéique ne sert que d'emballage.
* Remarque : si cette démonstration ne vous parait pas lumineuse, reportez vous à l'exercice (...et éventuellement à sa solution).
| relire ? |
