DISSECTION ULTIME DES GENES ET RECOMBINAISON IN VITRO
2 TECHNOLOGIES DE L'ADN RECOMBINANT
Les techniques d'étude de l'ADN sont devenues si performantes
qu'il
est actuellement courant d'isoler le segment d'ADN correspondant
à
n'importe quel gène spécifique.
La clé de voûte de ces technologies réside dans la
possibilité d'amplification de toute séquence
définie.
Cette amplification, le plus souvent par clonage du fragment d'ADN (un
clone
étant défini comme une population de cellules ou de
molécules
identiques à l'originale), permet d'obtenir des quantités
illimitées de molécules d'ADN de
séquence homogène.
2.1 CLONAGE DES GENES
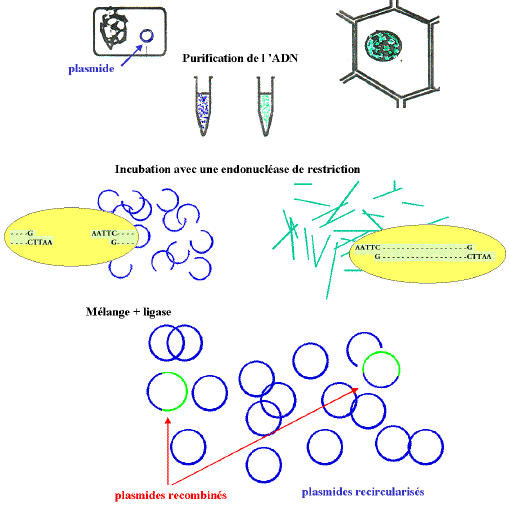
Un schéma ultra simplifié d'une expérience
typique de
clonage nécessite les éléments suivants :
l'ADN intéressant (à étudier ou pour ce
qu'il apporte) appelé également ADN passager, ADN
étranger ou ADN cible)
un vecteur de clonage (ou véhicule), dans l'exemple il
s'agit d'un plasmide c'est à dire un élément
génétique bactérien, extrachromosomique, capable
de réplication autonome.
une nucléase de restriction
une ligase
une cellule (procaryote ou eucaryote) pouvant servir
d'hôte biologique.
De nombreuses enzymes de restrictions génèrent des
extrémités cohésives et l'on peut insérer
des
fragments d'ADN digérés dans un vecteur présentant
les
mêmes extrémités c'est à dire traité
par
la même enzyme. Les nouvelles molécules sont ensuite
refermées
par une ligase. Cette recombinaison in vitro
conduit à une molécule "chimère" qui sera
transférée dans un hôte approprié par des
moyens
qui dépendent du choix de la cellule hôte. Après un
temps
de culture convenable on pourra obtenir des clones contenant chacun un
vecteur
recombiné, les clones différent entre eux par la nature
de
la séquence insérée dans le vecteur.
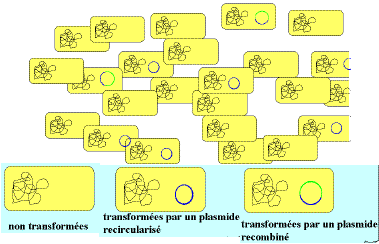
Comme le montre la figure ci-contre, après l'étape de
transformation génétique de l'hote, la population se
compose
en fait d'une grande quantité de cellules non
transformées
et parmi les bactéries transformées, beaucoup le sont par
un
vecteur recircularisé. Il faut donc pouvoir isoler rapidemant
les
clones recombinés et caractériser les
molécules
recombinées, c'est cette partie du travail, la plus
délicate,
qui a apporté énormément de renseignements sur la
structure
des gènes.
Le procédé de clonage lui même n'est qu'un moyen
pour
identifier et isoler des gènes rapidement.
2.2 LES VECTEURS DE CLONAGE
Comme on le voit, le vecteur joue un
rôle
essentiel dans toute expérience de clonage. On va donc
étudier
les véhicules avant de parler des passagers.
La plupart des fragments d'ADN étant incapable de
réplication
autonome, les vecteurs sont dérivés de réplicons
naturels
tels que les phages et les
plasmides.
Les plasmides sont des petites molécules circulaires d'ADN
double
brin, ils portent certains gènes fonctionnels (tels que des
gènes
de résistance à certains antibiotiques) et on les appelle
parfois
"les pièces de rechange génétiques" des
bactéries.
Les plasmides naturels ne sont pas utilisés tels quels car un
vecteur
doit répondre à un certain cahier des charges :
être aussi petit que possible car, dès que l'on
dépasse 15kb, l'efficacité de transformation de la
cellule hôte diminue rapidement.
être bien cartographié (gènes et sites de
restriction) l'idéal étant de connaître la
séquence complète.
il doit se répliquer facilement dans la cellule
hôte afin que de grandes quantités de molécules
recombinées puissent être préparées.
il doit posséder un marqueur de sélection qui
permette de distinguer les cellules transformées par le vecteur
des cellules non transformées
un vecteur idéal devrait de plus posséder un
marqueur qui permette de distinguer les clones transformés par
un vecteur recombiné de ceux transformés par ce vecteur
non recombiné.
il doit posséder le plus possible de sites de restriction
différents (mais chacun en un seul exemplaire) pour permettre
une grande souplesse dans le clonage de différents produits de
restriction.
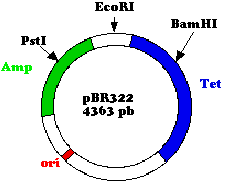
L'une des plus célèbres constructions est celle du
plasmide
pBR 322 (Bolivar et Rodriguez), de
très
nombreux vecteurs utilisés actuellement en sont
dérivés.
Le succès de ce vecteur tient à deux points :
il s'agit d'un plasmide dont la réplication est sous
contrôle "relâché" contrairement à beaucoup
d'autres qui sont sous contrôle "stringent" c'est à dire
que leur réplication est dépendante de la synthèse
protéique et de l'ADN polymérase III, on en trouve
généralement de 1 à 5 par cellule, les plasmides
sous contrçole "relaché" utilisent la polymérase I
et peuvent se répliquer en absence de synthèse
protéique, on en trouve de 30 à 50 copies par cellule ce
qui augmente l'amplification.
deux gènes de résistance, l'un à
l'ampicilline, l'autre à la tétracycline peuvent servir
de marqueurs pour sélectionner aisément les clones
transformés par des plasmides recombinés.
La figure ci-contre schématise ce vecteur de 4363 paires de
bases
qui contient des sites de restriction en un seul exemplaire (sites de
restriction
uniques pour plusieurs nucléases) dont certains sont
situés
dans les gènes de résistance.
L'insertion de la séquence à cloner dans l'un ou
l'autre de
ces gènes de résistance (au site BamHI dans le
gène
de résistance à la tetracycline, dans l'exemple ci
dessous)
l'inactive, ce qui offre un moyen très pratique d'identification
des
clones
transformés par des vecteurs recombinés selon le
génotype
:
clone
génotype
non transformé
ampS,tetS
transformé par un
vecteur recircularisé
ampR,tetR
transformé par un
vecteur recombiné
ampR,tetS
(*)
(*) dans
l'exemple
ci-dessus
D'autres vecteurs dérivés de celui-ci présentent
des
améliorations portant essentiellement sur la
facilité de criblage des clones
recombinés, sur la facilité
d'insertion de l'ADN étranger, sur
l'adaptation à l'utilisation
ultérieure de l'ADN cloné (cartographie,
séquençage, expression, étude de la
régulation
de l'expression...) et sur la taille des
segments
insérés.
Le bactériophage M13 par
exemple a été modifié pour en faire un vecteur, il
présente la particularité de posséder un
génome à ADN simple brin ce qui sera intéressant
pour le séquençage par exemple, après infection,
un brin complémentaire est synthétisé, il se
comporte donc comme un plasmide à l'intérieur de la
bactérie et pourra être manipulé comme tel.
* Remarque : N'oublions pas que les recombinaisons in vitro
utilisent les propriétés des nucléases de
restriction, le substrat est donc de l'ADN bicaténaire.
Le bactériophage lambda possède
une grande région de son génome non essentielle à
sa reproduction, elle pourra être éliminée et
remplacée par un ADN passager de l'ordre de kilopaires de bases
alors qu'un plasmide n'en tolère qu'une quinzaine.
Les cosmides combinent les
avantages de clonage des plasmides et augmentent la taille d'insertion
possible dans les phages lambda jusqu'à, une quarantaine de kilo
paires de bases.
Les BAC ou chromosomes
bactériens artificiels (Bacterial Artificial Chromosomes) sont
dérivés du facteur F de conjugaison (voir le chapitre
"Régulation de l'expression des gènes"). Ils
possèdent donc une origine de réplication et peuvent,
naturellement, intégrer des fragments d'ADN bactérien
allant jusqu'à 300kpb. Utilisés comme vecteurs, ils
permettent de cloner des fragments de 100-150 kpb et présentent
tous les avantages des vecteurs plasmidiques.
Les YAC ou chromosomes de levure
artificiels (Yeast Artificial Chromosomes) sont complétement
differents des prézcédents car dérivés
d'éléments eucaryotiques et seront hébergés
et reproduits par des cellules de levure. La construction repose sur
des séquences centromériques et
télomériques et une origine de réplication de
levure. Un site de clonage permet d'inserer des fragments d'ADN
étranger allant jusqu'à 2Mpb. Ces éléments
sont dupliqués et ségrègent à la mitose
comme des chromosomes. Ils représentent les seuls vecteurs
capables de réaliser des ensembles de clones chevauchant pouvant
couvrir l'ensemble d'un génome Eucaryote complexe.
Il faut donc choisir le vecteur en fonction des cribles dont on
disposera
pour repérer le clone intéressant et en fonction de la
nature
de l'ADN cloné c'est à dire du "passager".
2.3 CONSTRUCTION DE "BANQUES DE GENES"
Le vecteur étant choisi, la première étape est
celle
de la préparation de l'ADN à insérer. On va
distinguer
deux procédures, l'une conduisant à la réalisation
de
banques génomiques l'autre à
des
banques d'ADN complémentaire.
2.3.1 CAS DE LA CONSTRUCTION D'UNE BANQUE GENOMIQUE
L'ADN est préparé à partir du tissu le mieux
adapté,
puis découpé d'une façon la plus aléatoire
possible
(par cassures mécaniques ou par digestion incomplète par
une
nucléase de restriction). La construction d'une banque
génomique
implique le clonage de la totalité du génome, le seul
moyen
d'y parvenir est de partir de fragments chevauchants.
Tous les fragments de taille admissible par le vecteur choisi auront la
possibilité d'être clonés, tout le génome
pourra
être représenté. Le nombre minimum de clones
nécessaires pour qu'une séquence quelconque, appartenant
à
un génome de taille donnée soit présent dans la
banque
(qui, théoriquement, contient l'ensemble des séquences
d'ADN
de l'espèce considérée) peut se calculer par :
ln(1-P)
N = ---------------
L-X
ln 1- ------
M
N : nombre de clones constituant la banque (ou bibliothèque)
P : probabilité pour qu'une séquence donnée soit
présente dans la banque (une valeur de 0,99 est acceptable) L :
taille
moyenne des fragments insérés
X : taille de la séquence souhaitée
M : taille du génome
Quelle que soit la prétention de cette formule,
ceci
veut dire qu'une banque génomique réalisée dans un
vecteur
dérivé du phage lambda devra comprendre environ 40
000
clones différents pour un génome de drosophile et 800 000
pour
celui du maïs ou de l'homme. Ces banques sont donc très
lourdes
et dans bien des cas on constitue un autre type de banque.
2.3.2 CAS DE LA CONSTRUCTION D'UNE BANQUE D'ADN COMPLEMENTAIRES
Elles reposent sur une découverte qui a bouleversé le
dogme
central de la biologie moléculaire : pour certains virus dont le
matériel génétique est de l'ARN, il existe un flux
d'information de l'ARN vers l'ADN. Cette étape obligatoire pour
leur
reproduction est assurée par une enzyme particulière : la
transcriptase réverse. Elle permet une synthèse d'ADN
double
brin (c'est donc une ADN polymérase) à partir d'une
molécule
d'ARN. Elle représente également un outil très
utilisé
en génétique moléculaire. On peut en effet
synthétiser
in vitro des molécules d'ADN dont l'un des brins a une
séquence
parfaitement complémentaire de celle d'un ARN donné,
appelé
ADNc .
NB: in vitro, l'activité ribonucléasique de la
transcriptase
réverse est difficile à maîtriser et l'on utilise
souvent
d'autres stratégies enzymatiques pour synthétiser le
deuxième
brin d'ADN.
Les banques dites d'ADN complémentaire présentent les
avantages
suivants :
sachant que dans toute cellule, une très faible partie du
génome est transcrit en ARN, quel que soit la catégorie
cellulaire qui va servir de point de départ, la banque
nécessitera beaucoup moins de clones et l'enrichissement en
clones recherchés pourra être considérable si l'on
choisitbien un organe, son stade de développement...
on ne collectionne que des séquences codantes (sans
introns) qui pourront s'exprimer dans un hôte procaryote
même si elles sont d'origine eucaryote.
elles sont obligatoires pour cloner les gènes des virus
à ARN (la majorité des virus des végétaux).
2.4 PREPARATION DES VECTEURS RECOMBINES
Dans les
deux cas
: découpage mécanique d'ADN génomique ou
synthèse
d'ADN complémentaire, l'étape suivante est l'insertion
dans
un vecteur.
L'utilisation de nucléases de restriction produisant des
extrémités cohésives simplifie cette étape
mais
le découpage mécanique et la synthèse d'ADNc ne
procurent
pas de molécules à extrémités
cohésives.
Pour palier cet inconvénient, une technique encore largement
utilisée consiste à greffer des "queues"
homopolymériques
sur le vecteur et leurs complémentaires sur les fragments d'ADN.
On
utilise un site Pst I du vecteur car elle laisse des
extrémités
3' débordantes, ces extrémités vont être
allongées par un oligo dG grâce à la
transférase
terminale (enzyme qui polymérise, de façon non
spécifique,
des nucléotides à une extrémité 3' libre,
elle
n'utilise pas de modèle comme l'ADN polymérase). De la
même
manière, on greffe une queue oligo dC aux
extrémités
3' de l'ADN à insérer.
Après hybridation et ligation partielle, les cellules
hôtes
sont transformées, elles opèrent elles-mêmes une
réparation et la ligation recréant du même coup
deux
sites Pst I qui pourront être utilisés, après
amplification,
pour récupérer l'ADN cible à partir des clones
transformés.
Une autre méthode consiste à greffer, à l'aide
d'une
ligase, des oligonucléotides (4 ou 6 paires de
nucléotides)
représentant des sites de reconnaissance pour une
nucléase
de restriction aux extrémités franches des fragments
d'ADN
à cloner. Après ligation de ces petites molécules
adaptatrices, la digestion par l'enzyme appropriée va
créer
des extrémités cohésives. On aura la
précaution
d'avoir protégé au préalable d'éventuels
sites
internes de reconnaissance (par méthylation par exemple).
2.5 EXPLOITATION DES BANQUES
Plusieurs techniques éprouvées permettent de
construire des
banques. Il reste à les exploiter, même avec une banque
d'ADNc,
le nombre de clones est important et le tri représente la partie
la
plus délicate des expériences de clonage, il existe de
nombreuses
stratégies mais aucune n'est universelle.
* Remarque : les termes de cribler et
de
sélectionner n'ont pas la
même
signification: lors de la sélection, on
élimine tous les clones non
intéressants, le criblage permet de
repérer le clone
intéressant.
Le tri peut se faire par sélection lorsque le gène
intéressant confère un phénotype particulier
à
l'hôte tel qu'une résistance à un antibiotique
particulier
auxquel les cellules sauvages sont sensibles : la culture sur un milieu
contenant
cet antibiotique ne fera apparaître que les clones
transformés
contenant ce gène précis.
Le tri par criblage reste le plus courant.
La détection immunologique du produit du gène
représente
un crible intéressant si le gène recherché est
exprimé
dans la cellule transgénique ce qui n'est pas toujours le cas.
Le
criblage
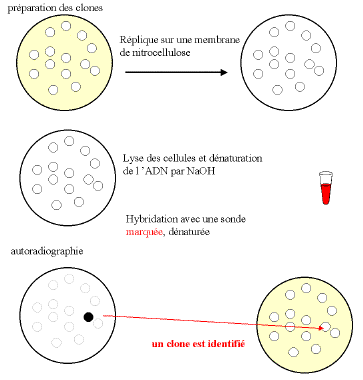
par hybridation de l'ADN des clones transformés avec une sonde
spécifique constitue une méthode de choix. L'hybridation
peut
se faire in situ : on effectue des répliques de clones
bactériens cultivés en boîte de Pétri sur
des
disques de nitrocellulose, après un temps de culture suffisant,
les
bactéries de ces répliques sont ensuite lysées par
la
soude qui, en même temps, dénature l'ADN, les
molécules
simple brin correspondantes se trouvent immobilisées à
l'emplacement de chaque clone. Après hybridation, avec la sonde
radioactive, l'autoradiographie révélera les clones
positifs.
Le problème est déplacé vers celui de
l'obtention de
la sonde spécifique, correspondant
au
gène recherché.
s'il s'agit d'un gène peu évolué, on peut
utiliser une sonde "hétérologue"
(en fait une séquence homologue mais provenant d'un autre
organisme) en comptant sur une homologie de séquence suffisante
pour s'hybrider dans des conditions qui ne donnent pas de faux
positifs.
lorsqu'une lignée cellulaire synthétise, à
un moment donné du développement, un messager
majoritaire, on peut tenter sa purification et réaliser un ADN
complémentaire qui, une fois cloné servira de sonde pour
une banque génomique.
si l'on connaît très bien le produit du
gène, c'est à dire la séquence, même
partielle, en acides aminés, on pourra construire des
oligonucléotides selon les codes possibles et "partir à
la pèche" avec ces sondes artificielles.
Dans les cas désespérés, c'est à
dire lorce que l'on ne possède pas de sonde et que l'on n'a
aucune idée du produit du gène, connu uniquement par la
manifestation phénotypique d'un allèle muté, il
reste d'autres solutions, par exemple, la "marche sur chromosome"
(chromosome walking) : Par des méthodes faisant appel à
des croisement traditionnels et à l'analyse mendélienne,
on va essayer d'associer des marqueurs RFLP au locus
considéré. L'idèal étant d'encadrer le
locus par des marqueurs distants de moins d'un centiMorgan. Le premier
marqueur constitue une sonde pour identifier un clone (parmi une banque
génomique) qui servira de départ. La cartographie de
restriction nous permettra d'identifier un segment situé
à une extrémité qui sera, à son tour
utilisé comme sonde pour cribler un clone chevauchant
et ainsi de suite jusqu'à un second marqueur connu.
La séquence recherchée se trouve obligatoirement parmi
les clones identifiés. Si l'un d'entre eux peut
complémenter un mutant (par transgénie restituant le
phénotype sauvage par exemple), c'est qu'il s'agit de la
séquence recherchée.
Le clonage biologique à l'aide d'un vecteur
approprié est applicable à n'importe quelle
séquence d'ADN (naturelle ou synthétique) et
représente un outil d'amplification de molécules
homogènes très puissant mais il ne s'agit que d'un outil.
Les technologies de l'ADN recombiné in vitro sont à la
base de la génétique moderne.
 polymérase III, on en trouve
généralement de 1 à 5 par cellule, les plasmides
sous contrçole "relaché" utilisent la polymérase I
et peuvent se répliquer en absence de synthèse
protéique, on en trouve de 30 à 50 copies par cellule ce
qui augmente l'amplification.
polymérase III, on en trouve
généralement de 1 à 5 par cellule, les plasmides
sous contrçole "relaché" utilisent la polymérase I
et peuvent se répliquer en absence de synthèse
protéique, on en trouve de 30 à 50 copies par cellule ce
qui augmente l'amplification. 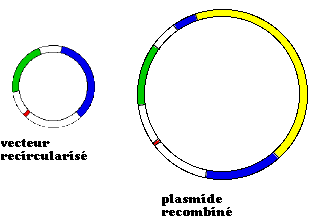 clones
transformés par des vecteurs recombinés selon le
génotype
:
clones
transformés par des vecteurs recombinés selon le
génotype
: Dans les
deux cas
: découpage mécanique d'ADN génomique ou
synthèse
d'ADN complémentaire, l'étape suivante est l'insertion
dans
un vecteur.
Dans les
deux cas
: découpage mécanique d'ADN génomique ou
synthèse
d'ADN complémentaire, l'étape suivante est l'insertion
dans
un vecteur.

 Le
criblage
par hybridation de l'ADN des clones transformés avec une sonde
spécifique constitue une méthode de choix. L'hybridation
peut
se faire in situ : on effectue des répliques de clones
bactériens cultivés en boîte de Pétri sur
des
disques de nitrocellulose, après un temps de culture suffisant,
les
bactéries de ces répliques sont ensuite lysées par
la
soude qui, en même temps, dénature l'ADN, les
molécules
simple brin correspondantes se trouvent immobilisées à
l'emplacement de chaque clone. Après hybridation, avec la sonde
radioactive, l'autoradiographie révélera les clones
positifs.
Le
criblage
par hybridation de l'ADN des clones transformés avec une sonde
spécifique constitue une méthode de choix. L'hybridation
peut
se faire in situ : on effectue des répliques de clones
bactériens cultivés en boîte de Pétri sur
des
disques de nitrocellulose, après un temps de culture suffisant,
les
bactéries de ces répliques sont ensuite lysées par
la
soude qui, en même temps, dénature l'ADN, les
molécules
simple brin correspondantes se trouvent immobilisées à
l'emplacement de chaque clone. Après hybridation, avec la sonde
radioactive, l'autoradiographie révélera les clones
positifs.
