2 DECOUVERTE DE LA FONCTION DU GENE
Une relation assez précise entre les unités d'information et la fonction
enzymatique a été établie par Beadle et Tatum vers 1941 à l'aide d'un champignon
ascomycète : Neurospora crassa dont le cycle biologique est représenté
ci-dessous.
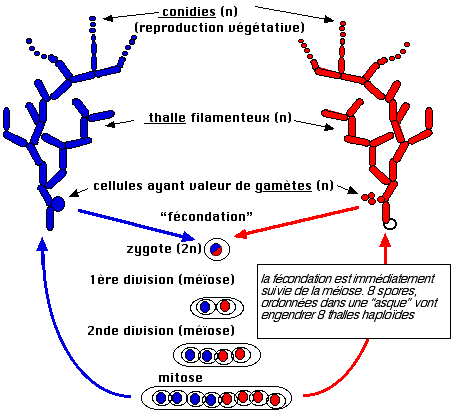 Cet organisme, bien que méiotique, est
supposé plus simple que le pois et la drosophile : la phase de croissance est
haploïde ce qui évite les complications liées aux
rapports allèliques, une phase de reproduction asexuée, par conidies
(bourgeonnements cellulaires uni ou oligo-nucléés) permet une mutagénèse en
masse (par irradiation par exemple) et l'obtention rapide de nombreux variants
Cet organisme, bien que méiotique, est
supposé plus simple que le pois et la drosophile : la phase de croissance est
haploïde ce qui évite les complications liées aux
rapports allèliques, une phase de reproduction asexuée, par conidies
(bourgeonnements cellulaires uni ou oligo-nucléés) permet une mutagénèse en
masse (par irradiation par exemple) et l'obtention rapide de nombreux variants
Ces champignons se cultivent aisément sur un
milieu défini très simple composé de sucre, de quelques sels et acides
inorganiques, d'un composé ammoniacal source d'azote et d'une vitamine : la
biotine. Les souches sauvages capables de croître sur ce milieu minimal sont
dites prototrophes, elles sont capables de réaliser
la synthèse de toutes les autres molécules nécessaires à leur métabolisme
(précurseurs des glucides, lipides, protéines, acides nucléiques et
macromolécules elles mêmes) à partir du milieu évoqué ci-dessus appelé milieu minimal. Une mutation peut très bien affecter une
des multiples fonctions du métabolisme de telle sorte que la souche devient
incapable de synthétiser un composé essentiel et donc de croître sur le milieu
minimal : la mutation est létale car, la cellule
étant haploïde, un locus homologue ne peut compenser ce défaut.
 Nous
allons définir un outil de travail indispensable : la mutation létale conditionnelle, en effet, la mutation n'est
létale que par rapport au milieu minimal utilisé, si l'on ajoute à ce milieu le
composé organique que la souche ne sait plus réaliser (on parle de supplémenter
le milieu), elle se développera comme une souche sauvage. Une souche auxotrophe est une souche ayant subi une telle mutation
nutritionnelle. Bien que de génotype muté, elle pourra, malgré tout, se
développer dans un milieu capable de complémenter l'effet de la mutation et
présentera alors un phénotype sauvage.
Nous
allons définir un outil de travail indispensable : la mutation létale conditionnelle, en effet, la mutation n'est
létale que par rapport au milieu minimal utilisé, si l'on ajoute à ce milieu le
composé organique que la souche ne sait plus réaliser (on parle de supplémenter
le milieu), elle se développera comme une souche sauvage. Une souche auxotrophe est une souche ayant subi une telle mutation
nutritionnelle. Bien que de génotype muté, elle pourra, malgré tout, se
développer dans un milieu capable de complémenter l'effet de la mutation et
présentera alors un phénotype sauvage.
Le protocole expérimental qui permet
d'obtenir rapidement de grandes collections de  mutants d'auxotrophie
est résumé ci-contre.
mutants d'auxotrophie
est résumé ci-contre.
Remarque : au cours de la mutagénèse par irradiation, celle-ci
n'induit pas de mutations précises, elle augmente considérablement la fréquence
des mutations mais d'une façon aléatoire et de très nombreuses mutations létales
passent inaperçues. C'est l'expérimentateur qui, par le jeu des milieux, choisit
la nature de l'auxotrophie et sélectionne les mutants correspondants.
La réalité de la mutation, c'est à dire la preuve d'une
altération héréditaire du matériel génétique est obtenue par croisement d'une
souche auxotrophe avec une souche sauvage de signe compatible. Chez Neurospora,
les ascospores restent ordonnées, dans l'asque, selon
l'orientation des fuseaux des deux divisions méiotiques et de la mitose
surnuméraire. Il est facile de déterminer individuellement les génotypes et de
montrer une répartition mendélienne de l'auxotrophie dans la descendance.
De très nombreux mutants auxotrophes, différents dans leurs exigences ont été
sélectionnés et le croisement entre ces mutants indique clairement que les
mutations portent sur des loci différents.
| On a donc une correspondance entre une mutation et
la disparition d'une fonction enzymatique nécessaire à l'accomplissement
d'une voie métabolique. |
Ces observations ont conduit à l'hypothèse célèbre dite "un gène, une enzyme"
qui, sous la forme "un gène une protéine" sera pleinement confirmée par
l'analyse ultérieure et indique la relation entre le génotype et le phénotype :
une protéine est un caractère phénotypique.
La richesse des collections de mutants d'auxotrophie devint rapidement telle
que l'on s'aperçut que des mutations entraînant le même phénotype (la même
auxotrophie), pouvaient se produire dans des gènes différents et il fallut
classer ces mutations.
Par exemple, Beadle et Tatum ont analysé une
collection de mutants exigents en arginine (mutants
arg-) et montré, par des tests de complémentation
adaptés à ce matériel que les différentes mutations conduisant à ce même
phénotype ne se situent pas dans le même cistron.
Test de complémentation :
la rencontre de deux thalles filamenteux différents peut entraîner une fusion
membranaire et l'apparition de cellules binuclées ou dicaryons. Dans de telles cellules, la présence de deux
noyaux de génotype différent dans un même cytoplasme peut permettre une
complémentation (la recombinaison est évidemment impossible).
Grâce à ce
test, il a été possible de réunir les différents mutants
arg- en groupes de complémentation ( argA, argB, argC
etc...) qui correspondent à des gènes différents.
Puisque la perte de
fonction de l'un quelconque de ces gènes entraîne le même phénotype :
auxotrophie arginine, il est clair que la synthèse de l'arginine nécessite
plusieurs unités d'information.
Beadle et Tatum ont précisé les exigences de
chacun de ces groupes en ajoutant au milieu des composés proches de l'arginine
et montré que ces exigences sont différentes pour chaque groupe de
complémentation..
Par exemple, les souches argH ne peuvent croître que sur
un milieu contenant de l'arginine (ce qui est normal puisqu'elle a été
sélectionnée en raison de cette déficience), par contre, la souche argG peut
croître en présence d'arginine mais pousse tout aussi bien si le milieu contient
de l'arginosuccinate.
La souche argF peut être complémentée par de
l'arginine, de l'arginosuccinate mais aussi de la citrulline et la souche argE
par l'un des trois composés précédents ou de l'ornithine.
Ceci cadre parfaitement avec l'hypothèse "un gène, une enzyme" la synthèse
des molécules biologiques se fait par une cascade d'actions enzymatiques (voie
métabolique) chaque étape est sous la dépendance d'un gène. Chaque gène est
susceptible d'être modifié par une mutation conduisant à un allèle nul, ce qui
entraîne la perte d'une fonction enzymatique.

La première expression phénotypique de chaque
gène est une protéine. C'est parce qu'ils gouvernent la production
d'enzymes spécifiques de façon régulée que les gènes sont responsables de
l'élaboration de l'ensemble du
phénotype. |