2 STRUCTURE ET PROPRIETES DES MOLECULES D'ADN
Watson et Crick quant à eux ont immédiatement compris la
portée de cette conclusion et leur proposition, en 1953, d'un modèle
moléculaire en double hélice maintenue par des liaisons
hydrogène entre des bases précises a marqué une autre
étape sensationnelle et décisive de la génétique
moderne. Les éléments de cette découverte sont
intéressants à rappeler car ils fournissent un bel exemple
de ce que permet l'intégration d'observations résultant de
disciplines différentes. Watson et Crick ont progressivement
élaboré leur modèle moléculaire à partir
d'images, souvent difficiles à interpréter, de diffraction
des rayons X par la molécule d'ADN laissant supposer une certaine
régularité et une certaine répétition dans cette
molécule très longue. Ils ont également tenu compte
des observations d'Erwin Chargaff portant sur la composition en bases
d'ADN
provenant de différentes sources. A l'époque, il n'était
pas question d'obtenir la séquence de ce polymère, mais il
était possible, après hydrolyse complète (c'est à
dire rupture de toutes les liaisons covalentes unissant les monomères
entre eux), de séparer ceux-ci par chromatographie sur papier.
On sépare ainsi quatre constituants : "A","T","G" et "C" (voir
ci dessous la nature de ces constituants), que l'on peut doser afin
d'évaluer leurs proportions respectives.
Le tableau II 1, présente des caractéristiques que Chargaff
a su interpréter : les purines (A et G) et les pyrimidines (C et T)
sont également représentées (50% de chaque), quelque
soit la source de l'ADN, la proportion d'adénine est la même
que celle de thymine et la proportion de guanine est la même que celle
de cytosine, par contre, le rapport A + T / G + C semble
caractéristique
de la source d'ADN.
| Organisme | A | T | G | C |
rapport A+T/G+C |
| E. coli D.pneumoniae Levure Rat Homme |
26,0 29,8 31,3 28,6 30,3 |
23,9 31,6 32,9 28,4 30,3 |
24,9 20,5 18,7 21,4 19,9 |
25,2 18,0 17,1 21,5 19,8 |
1,00 |
Tableau II.1
2.1 STRUCTURE DES ACIDES NUCLEIQUES
Les acides nucléiques sont des polymères de nucléotides de très grande taille (macromolécules).
Dans ce rappel très rapide on ne fera que souligner ce que les particularités structurales entraînent comme conséquences pratiques. Ainsi la composition détaillée des différents nucléotides est supposée connue mais il est bon de garder à l'esprit que leur enchaînement conduit effectivement à des molécules de très grandes tailles. Celles d'ADN se mesurent en centimètres pour la longueur avec un diamètre pour la double hélice de 20 Angstroem. Conséquence pratique : étant donné les moyens mécaniques mis en jeu dans les différentes étapes de purification de l'ADN, il est impossible d'obtenir des molécules intactes, on ne travaille qu'avec des morceaux. De plus, les cassures étant aléatoires, on travaille avec un mélange hétérogène de morceaux, c'est ce qui, pendant longtemps, à bloqué la progression de l'analyse fine du gène.
2.1.1 LES MONOMERES ET LEUR ENCHAINEMENT :
Le nucléotide est lui même composé de trois molécules :
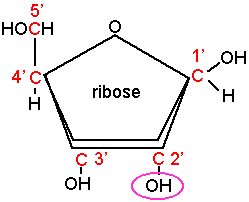
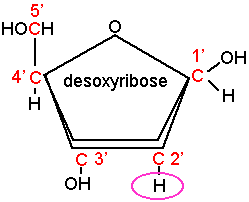
- un pentose sous forme cyclique (furane): le ribose en ce qui concerne l'acide ribonucléique (ARN) et le 2'désoxyribose pour l'acide désoxyribonucléique (ADN)
- une base organique est reliée au sucre en 1' : soit une base purique adénine (A) ou guanine (G) soit une base pyrimidique : cytosine (C), thymine (T) ou uracile (U), l'ensemble constitue un nucléoside
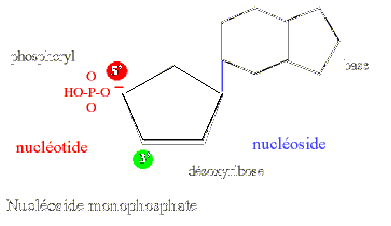
- un acide phosphorique vient estérifier une fonction alcool du sucre en 3' pour constituer un nucléotide.
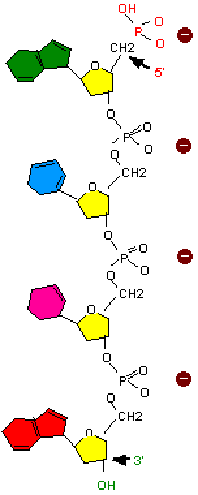
Les nucléotides précurseurs de la synthèse d'acides nucléiques sont sous forme triphosphate, deux phosphoryls (ß et ) seront éliminés au cours de la polymérisation.
La polymérisation s'effectue par l'estérification d'un alcool situé en 3' d'un nucléotide par le phosphate d'un autre nucléotide (la liaison covalente ainsi établie est dite liaison 3'-5' phosphodiester).
* Remarques pratiques :
1) En raison de leurs doubles liaisons conjuguées, les bases absorbent fortement la lumière ultraviolette. Il est donc très facile de doser les acides nucléiques en solution en mesurant la densité optique à 260 nm (maximum d'absorption).
2) En raison des groupes phosphoryls, les acides nucléiques sont chargés négativement, ils se comportent comme des polyanions et migrent vers l'anode lors d'une électrophorèse.
L'électrophorèse est actuellement l'outil de base de la génétique moléculaire car elle permet de séparer des fragments d'acides nucléiques en fonction de leur taille.
* remarque importante :
Les molécules se déplacent dans un champ électrique uniquement parce qu'elles sont chargées mais la migration nécessite un support . Sur le plan électrique, ce support (un gel poreux) n'intervient pas, par contre, sa porosité va déterminer, à charges égales, la vitesse de migration : des grosses molécules seront beaucoup plus freinées par le support que des petites. Cette propriété, même si l'explication reste intuitive, a un champ d'application considérable : la distance de migration est inversement proportionnelle au logarithme de la masse, ainsi, on va pouvoir, à l'aide de fragments de taille connue (échelle), "calibrer" les supports d'électrophorèse et déterminer, avec une précision qui dépend de la nature du gel utilisé, la taille des fragments d'ADN contenus dans un mélange..
Pour un acide nucléique donné, seules les bases distinguent les différents nucléotides. Les deux acides nucléiques sont des polymères à quatre monomères possibles symbolisés par A T G C pour l'ADN et A U G et C pour l'ARN.
Ces polymères ne sont pas ordonnés (de type ABCD ABCD ABCD ABCD...) mais séquencés ceci se traduit par une succession de nucléotides qui nous semble aléatoire. Cependant la séquence des monomères de tout acide nucléique est capitale pour la transmission et l'expression du matériel génétique. La séquence des nucléotides dans le polymère impose ce que l'on appelle la structure primaire des polynucléotides.
Le plus souvent, les molécules sont linéaires et possèdent donc une extrémité 5'phosphorylée et une extrémité 3'hydroxylée d'où la représentation simplifiée suivante :
5'P ......................................................................... 3'OH
2.1.2 STRUCTURES SECONDAIRES
La structure secondaire des acides nucléiques est imposée par l'appariement des bases c'est à dire la formation de liaisons hydrogène entre deux bases organiques.
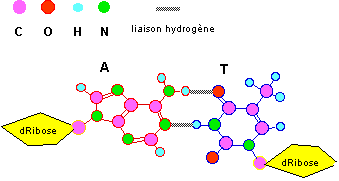
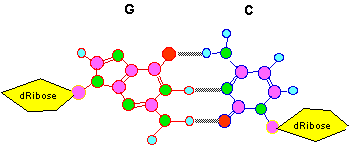
L'ADN est constitué de deux chaînes polynucléotidiques ainsi appariées. Etant donné que, pour des raisons d'encombrement stérique, deux paires de bases seulement sont stables A..T et G..C , la séquence d'un brin de cette molécule bicaténaire implique celle de l'autre.
Cette notion de complémentarité est fondamentale pour tout ce qui concerne le métabolisme de l'ADN in vivo et in vitro.
De plus, les deux bases d'une paire ne sont suffisamment proches l'une de l'autre et ne sont capables d'établir des ponts hydrogène que si les deux chaînes nucléotidiques sont antiparallèles soit :
5'P 3'OH
A C G T A T G C C C A T
A C
G C G C G C G
T G C A T A C G G G T A T G C G C G
C
G C
3'OH 5'P
Il en résulte, dans l'espace, une double hélice dont le pas de 34 A correspond à l'empilement de 10 paires de bases.
*Remarque : la figure précédente montre qu'il existe 3 liaisons hydrogène entre G et C et deux seulement entre A et T, ceci implique qu'un ADN riche en G-C sera plus stable qu'un ADN comportant plus de couples A-T.
Des molécules simple brin adoptent souvent, localement, des structures secondaires, selon le même principe d'appariement des bases.
C'est le cas de l'ARN : en raison des séquences soulignées dans la molécule ci-dessous:
5'P A U C G G C U U A G A C G A U G A A G C C G U C C C G G A A A 3'OH
elle peut prendre la structure secondaire
A
G A
U
G
A
C
G C
C G
A U
G
C
5'P
A U C G G C U U A C C G G A A A
3'OH
Enfin, parmi les structures particulières, il faut signaler celle des molécules circulaires : il n'y a pas d'extrémité 5' ni 3' (une liaison phosphodiester les relie). De telles structures (double ou simple brin), sont fréquentes chez les virus et des éléments génétiques particuliers tels que les plasmides, le "chromosome" bactérien est également une molécule circulaire
2.2 PROPRIETES TOPOLOGIQUES FONDAMENTALES DE L'ADN
Les liaisons hydrogène, relativement fragiles, peuvent être
détruites par chauffage ou par un pH élevé, dans ces
conditions les structures secondaires disparaissent Pour l'ADN, le
résultat
est la séparation complète des deux brins qui le composent
: il y a dénaturation de la
molécule.
En raison de la stricte complémentarité des bases, la
dénaturation est réversible, les
deux brins peuvent, dans des conditions appropriées de température
et de force ionique, rétablir des liaisons hydrogènes entre
leurs bases et reprendre la configuration en double hélice
d'origine.
Il faut bien comprendre que ce phénomène ne dépend que
de le complémentarité de deux séquences nucléotidiques
et non de l'origine de chaque brin.
Dans le cas de renaturation, on a formation de molécules "homoduplex"
c'est à dire de structures doubles, chaque partenaire ayant une même
origine. On peut très bien mélanger des ADN dénaturés
d'origines différentes s'il y a des complémentarités
de séquences des "hétéroduplex" c'est à dire
des molécules dont les deux brins sont d'origine différente
pourront se former. Enfin, toujours par complémentarité de
séquences, on peut apparier un ARN avec un brin d'ADN (il existe une
possibilité de liaison A..U) on obtient alors une molécule
"hybride". (voir ci-dessous)
Cette propriété (appariement de séquences complémentaires) représente un outil puissant dans la reconnaissance de séquences précises à l'aide de petits fragments polynucléotidiques appelés "sondes" car, dans un ensemble complexe de séquences différentes, elles sont capables de trouver celle qui leur correspond, de s'y hybrider La détection de ces hybrides sera facilitée par l'utilisation de sondes "marquées".
Exemples :
molécule native :
A A T G C C G T C A C T T T A G C T A T
A
T T A C G G C A G T G A A A T C G A T A
T
molécule dénaturée :
A A T G C C G T C A C T T T A G C T A T A
T T A C G G C A G T G A A A T C G A T A T
homoduplex (les deux brins renaturés sont de même origine :
A A T G C C G T C A C T T T
A
G C T A T A
T T A C G G C A G T G A A A T C G A T A T
heteroduplex (les dux brins d'ADN renaturés sont d'origine différente) :
A A T G C C G T C A C T T T A G C
T
A T A
T G A A A T C G A T A T G G G A
C
hybride (appariement d'un brin d'ADN et d'un brin d'ADN) :
A A T G C C G T C A C T T T A G C
T
A T A
U U A C G G
C
A G U G A A A U C G A U A U
sondes :
A A T G C C G T C A C T T T
A
G C T A T A
A C G G C A
ou
A A T G C C G T
C
A C T T T A G C T A T A
T C G A T
| relire ? |

© Université
de TOURS - GÉNET
Document modifié le
22 mars, 2010
![]()