L'étude de la recombinaison dans un croisement F1 X F1 est particulière.
Elle est possible, si l 'on reprend les deux souches que nous avons étudiées.
Dans ce cas, les individus de référence proviennent de gamètes
femelles parentaux ou recombinés et ne peuvent donc pas être
utilisés pour étudier la recombinaison.
femelles X mâles
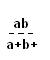 X
X
La fréquence de recombinés peut être estimée
par l 'observation des autres descendants car les individus a/a et b/b
représentent la moitié des recombinés. La fréquence
de recombinés est donc :
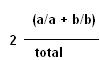
En revanche, si le croisement d 'origine est P1 : ab+ / ab+ X
P2 : a+b / a+b, l 'étude de la recombinaison n'est pas possible.
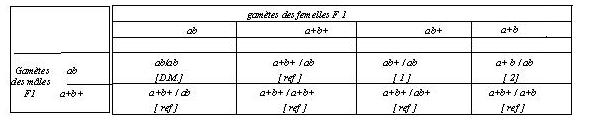
En effet, la F1 est alors ab+ / a+b et le tableau de rencontre des gamètes
qui en découle est:

Les individus correspondant à des gamètes parentaux ou
remaniés ne peuvent alors être distingués. On
remarquera qu 'il n 'y a pas d 'individus [double mutant] dans cette descendance.
Chez la levure : lorsque la distance
physique est très grande, nous avons vu que l 'on peut atteindre
une fréquence de recombinés de 0,50. Cela laisse persister
un doute lorsqu 'on observe une telle fréquence. En effet deux
hypothèses restent possibles : soit les gènes sont sur
des chromosomes indépendants, soit ils sont sur le même
chromosome mais très éloignés .
Chez la drosophile: l'absence de crossing-over chez
le mâle
permet de conclure très facilement, par la simple analyse qualitative
des individus [ double mutant ] dans le cas de ce croisement F1 X F1 :
s 'ils existent , les gènes sont sur des chromosomes différents ;
s 'ils sont absents , les gènes sont liés.