Réseau
GÉNET
Les outils
de
génétique
moléculaire
Les techniques liées aux acides nucléiques
Préparation des acides nucléiques
| Synthèse in vitro
Première manipulation génétique Les séries d'expériences réalisées par Kornberg et son groupe préfigurent la génétique moléculaire moderne et méritent que l'on s'y arrête. Le mode semi-conservatif de la synthèse de l'ADN implique les éléments suivants :
Polymérisation de desoxyribonucléotides in vitro Pour purifier et étudier cette enzyme, Kornberg a mis au point un système de synthèse in vitro à partir d'extraits d'abord assez grossiers d'E. coli. Comment évaluer de tels systèmes ? Comment prouver qu'une synthèse a bien lieu in vitro ? Comment distinguer l'ADN néosynthétisé de celui qui est obligatoirement présent dans l'extrait comme modèle ? Kornberg va lui aussi faire appel à des marqueurs isotopiques : des nucléotides comportant des phosphores 32 radioactifs (32P), si une synthèse a lieu, elle fera appel à ces précurseurs radioactifs et le polymère résultant sera "marqué", sera radioactif et facilement repérable. D'après l'analyse des nucléotides libres présents dans le cytoplasme, Kornberg décide de choisir des nucléotides triphosphorylés en 5' alors que les constituants de l'ADN sont monophosphorylés et que bien souvent, l'hydrolyse de polynucléotides produit des mononucléotides phosphorylés en 3' ! On verra que sans cette décision, l'expérience était vouée à l'échec : la cellule utilise effectivement des nucléotides 5' triphosphorylés et l'énergie fournie par la libération du pyrophosphate. *Remarque concernant l'utilisation de précurseurs radioactifs Dans son mélange réactionnel, Kornberg dispose d'ADN modèle, de précurseurs naturels, de l'ADN polymérase active (il l'espère), auquel il ajoute des précurseurs radioactifs. Après la réaction, la radioactivité se trouvera partagée entre l'ADN éventuellement synthétisé in vitro (en incorporant des monomères marqués) et l'excédent de précurseurs qui n'ont pas été incorporés. Il est donc essentiel d'éliminer tous ces précurseurs libres pour attribuer de la radioactivité à une macromolécule. En pratique, les macromolécules sont précipitées par adjonction d'un acide organique et les petites molécules "acido-solubles" (y compris les précurseurs radioactifs) sont éliminées par centrifugation. Le culot, après plusieurs lavages, contient les macromolécules (y compris l'ADN) débarrassées de tout précurseur non incorporé dans la chaîne. En suivant la stratégie exposée et ses contraintes, Kornberg fut capable de trouver quelque radioactivité dans des fractions acido-précipitables. Radioactivité qui, à l'époque ne dépassait guère le seuil de confiance des compteurs, mais Kornberg y croyait ! Plusieurs équipes, partant de kilogrammes de pâte d'E.coli, à l'aide de méthodes d'analyse biochimique classiques de nos jours mais que l'on découvrait à l'époque, ont peu à peu concentré l'activité de l'ADN polymérase jusqu'à purifier cette enzyme qui fut nommée "polymérase de Kornberg". Le bilan (provisoire) de ces expériences est le suivant : 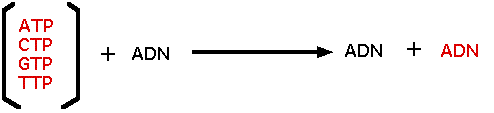
|
© Université de Tours - 21 mai 2002