Les Biotechnologies et l'agriculture : Techniques de base et applications

SOMMAIRE
|
|
Partie I: L'amélioration des plantes en quelques étapes clefs |
|
SOMMAIRE
|
|
Partie I: L'amélioration des plantes en quelques étapes clefs |
|

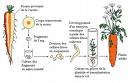
C'est la méthode empirique la plus ancienne. Elle a prévalu jusqu'à la fin du XIXe siècle, avant que l'on ne tire les conséquences des travaux de Mendel sur l'hérédité.
Cette méthode consiste à choisir les plantes qui semblent les plus intéressantes dans une population et à utiliser leurs graines comme semences pour la culture suivante. L'opération est répétée de génération en génération, ce qui permet d'améliorer progressivement les performances de la culture.
Entre deux étapes de sélection, les recombinaisons génétiques se font naturellement, sans aucun contrôle humain. Les plantes sélectionnées ne sont par conséquent ni identiques à celles de la génération précédente ni identiques entre elles. Elles ne constituent donc pas une variété.
Dans son principe, la sélection récurrente se rapproche de la sélection massale dans la mesure où l'objectif est d'améliorer la valeur moyenne d'une population en sélectionnant de génération en génération les meilleurs individus de cette population. La différence avec la sélection massale réside dans le fait que les croisements sont cette fois-ci contrôlés, et que la sélection récurrente conduit à la sélection de lignées ou d'hybrides.
![]() Pour en savoir plus sur la sélection récurrente;
Pour en savoir plus sur la sélection récurrente;
• La sélection généalogique
Cette méthode, aussi appelée filiation en autofécondation, se caractérise par le fait que la sélection des lignées a lieu pendant leur fixation à l'état homozygote.
|
• La méthode bulk
La méthode bulk est également appelée sélection généalogique différée car dans cette méthode, la sélection a lieu après fixation des lignées.
![]() Pour en savoir plus sur la sélection bulk
Pour en savoir plus sur la sélection bulk
• La sélection par filiation monograine
Cette méthode, aussi appelée Single Seed Descent (SSD), ressemble à la méthode bulk dans la mesure où la sélection des lignées intervient après leur fixation. La différence avec la méthode bulk réside dans le fait que l'on étudie ici la descendance de graines prises individuellement à chaque étape d'autofécondation.
![]() Pour en savoir plus sur la sélection SSD
Pour en savoir plus sur la sélection SSD
. • Le rétrocroisement
Le rétrocroisement, ou back-cross, est utilisé pour introduire un caractère intéressant, comme la résistance à une maladie par exemple, dans une lignées de bonne valeur agronomique.
Cette lignée est croisée avec une autre lignée : une plante sauvage apparentée ou une plante appartenant à un espèce proche qui possède le caractère d'intérêt. Après avoir vérifié que l'hybride possède ce caractère, on procède à une série de croisements avec la lignée d'intérêt agronomique (rétrocroisements), en sélectionnant à chaque étape les plantes qui portent le caractère d'intérêt.
Les rétrocroisements sont répétés pendant 6 à 8 générations, ce qui permet d'obtenir une lignée extrêmement proche de la lignée initiale mais qui possède en plus le caractère que l'on voulait introduire.
![]() Pour en savoir plus sur la sélection back cross
Pour en savoir plus sur la sélection back cross
• Les hybrides et l'hétérosis
La création d'hybrides se fait en deux étapes. La première consiste à créer et à sélectionner deux lignées qui seront ensuite croisées pour donner naissance à l'hybride.
La sélection des lignées nécessite l'utilisation d'un testeur : la lignée candidate à la sélection est croisée avec une autre lignée, le testeur, et l'étude de la descendance du croisement permet d'évaluer la valeur en croisement de la lignée candidate.
La lignée candidate peut être obtenue par l'une des méthodes présentées ci-dessous. On peut aussi utiliser des plantes issues d'un croisement et d'une autofécondation, qui ne sont donc pas fixées. On réalise alors 6 à 7 cycles successifs d'autofécondation avec à chaque fois un croisement avec le testeur et une sélection des meilleures lignées, ce qui permet d'obtenir des lignées fixées sélectionnées sur leur valeur en croisement.
L'hybride est ensuite obtenu en croisant l'une des lignées sélectionnées avec le testeur.A la différence d'une lignée, l'hybride est hétérozygote pour un grand nombre de gènes.
Les performances agronomiques de l'hybride sont en général meilleures que celles des deux parents : c'est le phénomène d'hétérosis, qui justifie l'utilisation d'une méthode de sélection qui est par ailleurs complexe et coûteuse. Cette méthode est utilisée pour des espèces telles que le maïs, le tournesol ou le colza. Le gain de rendement du fait de l'hétérosis est par exemple estimé à au moins 20% chez le colza.
L'amélioration des plantes a pour but de créer de nouvelles variétés à partir de la diversité existante. Elle consiste à croiser deux plantes choisies pour leurs caractères intéressants. Par le choix des meilleures plantes dans la descendance, les sélectionneurs aboutissent après un long travail d'épurations successives à la création d'une nouvelle variété.
Les objectifs de la sélection sont nombreux. Généralement, le premier critère évoqué est la productivité. 
La productivité dépend de nombreux facteurs. Elle peut être le résultat de la réduction des facteurs qui limitent le rendement, mais le potentiel de productivité peut également être accru par une amélioration de la physiologie des plantes : augmentation de l'activité photosynthétique, meilleure élaboration, migration et répartition des éléments constitutifs des réserves de la plante (grains, racines...). Les espèces végétales sont également plus ou moins plastiques. Certaines plantes comme le blé nécessitent une adaptation variétale importante aux conditions de sol et de climat. Pour les agriculteurs, l'un des facteurs les plus importants est la résistance aux maladies et aux parasites. Ceci joue non seulement sur le rendement, mais aussi sur le revenu de l'agriculteur.
De nouveaux objectifs ont été donnés à la sélection au fur et à mesure qu'évoluaient les demandes de l'industrie et de la société (par exemple: agriculture respectueuse de l'environnement, production de matières premières pour l'industrie chimique, production de molécules à usage pharmaceutique).


La vrai réforme de l'agriculture a été le passage d'un modèle d'agriculture familiale vivrière à un modèle d'agriculture qui nourrit "les villes". Au départ les agriculteurs produisaient pour nourrir leur famille avec une récolte qui s'étalait sur plusieurs mois. On mangeait au fur et à mesure les produits qui étaient à maturité. A partir de la fin du XIXe siècle, la production agricole a eu de plus en plus pour objectif de nourrir les populations urbaines alors même que le nombre d'agriculteurs diminuait.
Concilier augmentation de la production et diminution du nombre d'agriculteurs a nécessité un changement de production pour augmenter l'efficacité du travail (avec par exemple, le développement du machinisme agricole). Il a fallu notamment raccourcir le temps consacré à la récolte en faisant en sorte que l'agriculteur ne passe qu'une fois dans son champ.
Concentrer la récolte sur quelques jours par an sans diminuer la production implique que toutes les plantes du champ arrivent à maturité en même temps. Cette synchronisation n'a pu être obtenue qu'avec l'utilisation systématique des hybrides F1.
La synchronisation de la maturation pour permettre un ramassage mécanisé complet en un seul passage est le critère privilégié pendant la sélection qui conduit aux hybrides F1. En revanche, la production totale sur une année n'est pas très différente de celle du système antérieur où la récolte s'étalait sur plusieurs mois.

La révolution verte a été initiée en Inde à partir de 1966 et s'est ensuite rapidement propagée en Asie et Amérique du Sud. Le principe a été de développer un système d'agriculture totalement "collectiviste" basé sur des lignées qui tirent un profit maximum des engrais et de l'irrigation (par exemple des variétés à tige courte qui peuvent ainsi supporter sans casser le épis plus gros qui sont obtenus grâce aux apports d'engrais).
La révolution verte ne se limite pas à la création de variétés. Elle inclut un système cohérent de production: formation des agriculteurs ; aide financière pour l'accès aux produits phytosanitaires ; garantie des prix d'achats des récoltes.
Devant les résultats spectaculaires obtenus en Inde, ce système a été adopté avec succès dans la plupart des pays du tiers-monde. Sa principale limite est qu'il nécessite une abondante irrigation. Il ne constitue donc pas une solution durable dans de nombreuses régions du globe.

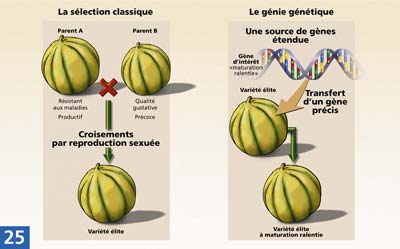
L'amélioration des plantes donne d'excellents résultats. Des schémas de sélection adaptés à chaque espèce ont été élaborés et optimisés. Ainsi, pour l'ensemble des espèces cultivées, des progrès importants ont pu être réalisés (productivité, adaptation au milieu, qualité technologique...). Cependant, cette voie se heurte à trois limites inhérentes à la reproduction sexuée : l'incompatibilité, l'imprécision et le temps. Les biotechnologies apportent de nouvelles techniques à la sélection classique : pour faciliter les croisements interspécifiques, pour maîtriser les transferts de gènes, pour créer rapidement des lignées pures.
Les sélectionneurs cherchent à réaliser des croisements interspécifiques (entre plantes d'espèces différentes mais génétiquement proches), afin d'augmenter les ressources en caractères favorables.
 Tomate de type cocktail à la couleur très originale : la tomate "bleue". Variété obtenue par l'OSU (Oregon State University), aux Etats-Unis. Cette tomate contient dans son épiderme un pigment bleu habituellement absent chez les tomates : les anthocyanes. Obtenu par croisement interspécifique avec une solanacée sauvage péruvienne riche en anthocyanes, cette tomate a donc une peau bleue. La chair est rouge foncé.
Tomate de type cocktail à la couleur très originale : la tomate "bleue". Variété obtenue par l'OSU (Oregon State University), aux Etats-Unis. Cette tomate contient dans son épiderme un pigment bleu habituellement absent chez les tomates : les anthocyanes. Obtenu par croisement interspécifique avec une solanacée sauvage péruvienne riche en anthocyanes, cette tomate a donc une peau bleue. La chair est rouge foncé.
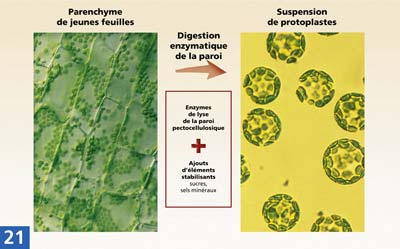
La création d'hybrides interspécifiques se heurte généralement à un obstacle biologique : absence de fécondation, avortement de l'embryon, ou obtention d'un descendant stérile. Le problème peut être résolu en ayant recours aux techniques de sauvetage d'embryons, de fusion de protoplastes et de transgénèse.
Lorsque le sélectionneur fait un croisement, il brasse un très grand nombre de caractères, aussi bien ceux qu'il désire introduire dans la nouvelle variété, que des caractères indésirables. Pour éliminer ces derniers, il doit ensuite procéder à de longues années de sélection
Le problème est en grande partie supprimé grâce à la transgénèse car elle permet d'introduire directement le gène intéressant dans une variété donnée sans en modifier le fond génétique. Cette technique n'est accessible que grâce à la connaissance du génome, à la réalisation de cartes génétiques, et à l'utilisation de marqueurs moléculaires.

La sélection est un processus de longue haleine. Il faut compter 5 à 15 ans, selon les espèces, pour créer une nouvelle variété et la mettre sur le marché.
Les techniques d'haplodiploïdisation et de culture d'embryons permettent de raccourcir la durée des cycles de sélection, en diminuant le temps nécessaire à la fixation et à la multiplication des génotypes intéressants.
Le clonage
On sait obtenir en nombre indéfini des copies végétatives à l'identique d'un individu: on prend par exemple des cellules de méristème et on les fait régénérer in vitro. L'intérêt évident est la multiplication en millions de copies d'une plante exceptionnelle, même si celle-ci est fortement hétérozygote.
Ces vitroplantes sont actuellement très répandues. C'est par exemple le cas de 80% des fraises consommées, de la quasi totalité des plantes vertes, des oeillets, d'une partie des rosiers, des plants souches de luzerne, de carotte ou de betterave.
L'hapoïdisation
Au lieu de partir de cellules de méristème, on peut partir de cellules sexuelles (donc haploïdes) pour régénérer une plante. C'est l'androgénése (on part de gamètes mâles) ou la gynogénèse (on part de gamètes femelles).
Un traitements chimique permet de retourner à l'état diploïde qui est l'état fertile. Comme cette transformation est obtenue par doublement du stock de chromosome, la plante résultante est homozygote. On obtient ainsi directement une lignée pure, ce qui représente un gain considérable par rapport aux méthodes de sélection traditionnelles.
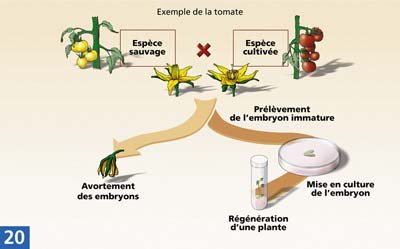
Le sauvetage d'embryons
Lors de croisements interspécifiques, des barrières naturelles empêchent le développement complet de l'embryon. Pour remédier à cette situation, on pratique après fécondation un prélèvement précoce des embryons pour les mettre en culture sur un milieu artificiel nutritif. Cette technique de culture in vitro est appelée sauvetage d'embryons interspécifiques.
Il est utilisé par exemple chez la tomate pour introduire dans l'espèce cultivée ( Lycopersicon esculentum) des gènes de résistance aux maladies présent dans l'espèce sauvage ( Lycoparium peruvianum). L'embryon est prélevé avant la phase de maturation de la graine. Il est ensuite transplanté et cultivé sur un milieu artificiel riche en sucre, permettant la régénération d'une plante nouvelle.
L'hybride obtenu est souvent croisé par le parent de l'espèce cultivée (rétrocroisement) et sa descendance est sélectionnée, pour fixer des caractères nouveaux et intéressants, tout en éliminant les caractères indésirables issus de l'espèce sauvage.
Pour en savoir plus sur le sauvetage d'embryons
La fusion de protoplastes
Le génie génétique, la transgénèse et les OGM
Le génie génétique désigne l'ensemble des techniques permettant d'introduire et de faire exprimer dans un organisme vivant un ou des gènes provenant de n'importe quel autre organisme. Les organismes ainsi obtenus sont dits Organismes Génétiquement Modifiés (OGM). On distingue les techniques de biologie moléculaire qui permettent de préparer les séquences d'ADN qui seront introduites, on parle de construction génétique, et les techniques de transgénèse qui permettent de transférer le gène.
Plusieurs découvertes scientifiques ont permis d'aboutir à l'obtention de la première plante transgénique en 1983. Notamment, la compréhension des mécanismes reponsables de la galle du collet, maladie connue depuis l'antiquité, a mis en évidence un transfert génétique naturel. Ce mécanisme de transfert est à l'origine des techniques de transformation génétique utilisées aujourd'hui.
Les deux principales caractéristiques du génie génétique en comparaison de la sélection classique basée sur la reproduction sexuée sont
 |
|
Il est important de noter que ces biotechnologies ne se substituent pas aux étapes classiques mais elles leur donnent plus de rapidité, plus d'envergure, plus de sécurité. Elles contribuent à une évolution accélérée de l'agriculture ce qui pose un certain nombre de questions que l'on abordera dans la partie débat discussion.
On peut citer à titre d'exemple de croisements interspécifiques ayant donné des hybrides d'intérêt commercial:







La tomate cultivée ![]() Lycopersicon esculentum possède une variabilité génétique faible. En revanche, les tomates sauvages possèdent de nombreux gènes de résistance aux maladies notamment l'espèce Lycoparium peruvianum. Les barrières d'incompatibilité avec les espèces sauvages génétiquement les plus éloignées de la tomate cultivée ont été contournées grâce au sauvetage d'embryons.
Lycopersicon esculentum possède une variabilité génétique faible. En revanche, les tomates sauvages possèdent de nombreux gènes de résistance aux maladies notamment l'espèce Lycoparium peruvianum. Les barrières d'incompatibilité avec les espèces sauvages génétiquement les plus éloignées de la tomate cultivée ont été contournées grâce au sauvetage d'embryons.
La courgette ![]() est une espèce fortement attaquée par l'oïdium, les virus de la mosaïque du concombre et de la mosaïque de la pastèque. Les variétés cultivées, Cucurbita pepo, sont insuffisamment résistantes à ces parasites. Des croisements avec des espèces sauvages américaines résistantes, Cucurbita okechobeenis et Cucurbita ecuadorensis, ont été réalisés grâce au sauvetage d'embryons immatures, rendant ainsi possible l'amélioration de la résistance aux maladies des variétés cultivées.
est une espèce fortement attaquée par l'oïdium, les virus de la mosaïque du concombre et de la mosaïque de la pastèque. Les variétés cultivées, Cucurbita pepo, sont insuffisamment résistantes à ces parasites. Des croisements avec des espèces sauvages américaines résistantes, Cucurbita okechobeenis et Cucurbita ecuadorensis, ont été réalisés grâce au sauvetage d'embryons immatures, rendant ainsi possible l'amélioration de la résistance aux maladies des variétés cultivées.
La laitue ![]() Lactuca sativa, est une espèce sensible au mildiou. Le mildiou de la laitue (Bremia lactucae) est la maladie la plus redoutée sur cette espèce, notamment en culture biologique d'hiver sous abris. Le croisement avec deux espèces sauvages, Lactuca virosa et Lactuca saligna, possédant des gènes de résistance intéressants est difficile à réaliser. Le sauvetage d'embryons interspécifiques a permis d'augmenter considérablement la réussite de ces croisements.
Lactuca sativa, est une espèce sensible au mildiou. Le mildiou de la laitue (Bremia lactucae) est la maladie la plus redoutée sur cette espèce, notamment en culture biologique d'hiver sous abris. Le croisement avec deux espèces sauvages, Lactuca virosa et Lactuca saligna, possédant des gènes de résistance intéressants est difficile à réaliser. Le sauvetage d'embryons interspécifiques a permis d'augmenter considérablement la réussite de ces croisements.
Puisqu'elle permet d'introduire un gène d'origine quelconque dans une plante donnée, la transgénèse peut potentiellement apportée des solutions à des problèmes très variés touchant aussi bien l'agronomie, l'amélioration des qualités alimentaires, la santé, l'industrie ou l'environnement.
Pourtant, les OGM commercialisés jusqu'ici visent un marché plus étroit: l'utilisation des herbicides chez le colza, le maïs et le soja et la lutte contre les insectes chez le coton. Le marché mondial correspondant est très important (les herbicides jouent un rôle important dans la culture du colza, du maïs et du soja; la culture du coton représente un quart de la consommation d'insecticides) et la surface mondiale cultivée en OGM augmente de 12% par an.
Il faut aussi noter la commercialisation récente de légumes OGM aux Etats-Unis et en Chine (papaye, poivron, tomate).
On notera aussi qu'en Mars 2010 la comission européenne à Bruxelles a donné son accord pour la culture d'une pomme de terre OGM en Europe. Il s'agit de la variété AMFLORA développée par BASF et autorisée pour un usage industriel (donc non alimentaire). L'amidon est composé d'amylose et d'amylopectine. S'ils ont tous les deux des vertus nutritionnelles, seule l'amylopectine est utile dans l'industrie. Jusqu'alors, cette séparation de l'amylose et de l'amylopectine était coûteuse. La pomme de terre de BASF, par une modification génétique qui inhibe la synthèse d'amylose, produit de l'amylopectine pure. La Commission européenne souligne donc que cette pomme de terre génétiquement modifiée «contribue à optimaliser le processus de production et à économiser des matières premières, de l'énergie, de l'eau et des substances pétrochimiques».
La réussite commerciale des OGM pour les principales plantes de grande culture n'empêche pas les débats. Ceux-ci sont présentés dans la partie débat-discussion. Ils sont plus faciles à comprendre lorsqu'on a en mémoire les contraintes économiques qui pèsent sur l'agriculture (voir la présentation du contexte agricole français) et la pression pour réduire dans les prochaines années l'usage des produits phytosanitaires (voir rapport INRA sur pesticides).

© Université
de TOURS - GÉNET
Document modifié le
26 mai, 2010
![]()
![]()
![]()